LUYOR-3109高強(qiáng)度紫外催化光源促銷
LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學(xué)透鏡,輸出紫外線強(qiáng)度高,...
2024-08-08


LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學(xué)透鏡,輸出紫外線強(qiáng)度高,...
2024-08-08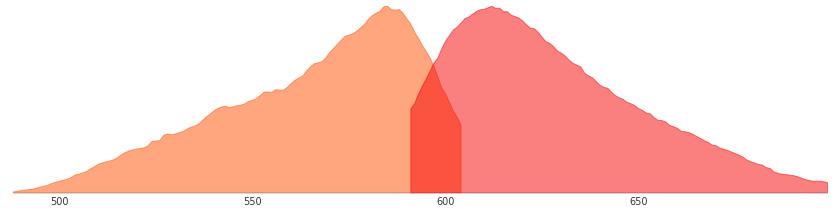
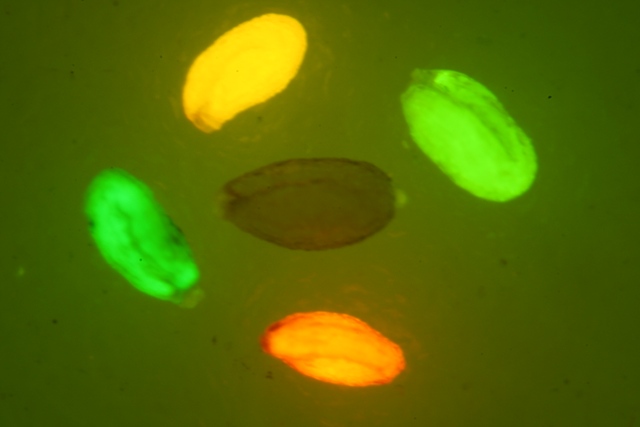
KillerRed是個(gè)完全由基因編碼的光毒性紅色熒光蛋白,可接受綠色光照(540~580nm)生成活...
2024-04-29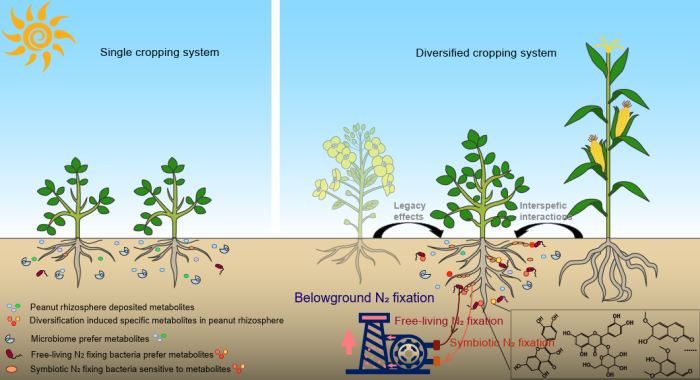
中國(guó)科學(xué)院南京土壤研究所彭新華研究員團(tuán)隊(duì)陳晏副研究員在農(nóng)田長(zhǎng)期多樣化種植下,種間植物根際對(duì)話調(diào)控土壤...
2024-04-07咨詢熱線
021-64195798作者:生命科學(xué)事業(yè)部時(shí)間:2019-12-14 20:06:48瀏覽3022 次
真核生物啟動(dòng)子和增強(qiáng)子是由若干可以區(qū)分的DNA序列組成的,由于它們和特定的功能基因連鎖在一起,因此稱為順式作用元件。真核生物轉(zhuǎn)錄調(diào)控大多是通過(guò)順式作用元件和反式作用因子復(fù)雜的相互作用而實(shí)現(xiàn)的,下面介紹的是與順式作用成分專一性結(jié)合的一些轉(zhuǎn)錄因子。一般認(rèn)為,如果某個(gè)蛋白是體外轉(zhuǎn)錄系統(tǒng)中起始RNA合成所必需的,它就是轉(zhuǎn)錄復(fù)合體的一部分。
真核基因組的一般構(gòu)造特點(diǎn)
①在真核細(xì)胞中,一條成熟的mRNA鏈只能翻譯出一條多肽鏈,不存在原核生物中常見(jiàn)的多基因操縱子形式。
②真核細(xì)胞DNA都與組蛋白和大量非組蛋白相結(jié)合,只有一小部分DNA是裸露的。
③高等真核細(xì)胞DNA中很大部分是不轉(zhuǎn)錄的,大部分真核細(xì)胞的基因中間還存在不被翻譯的內(nèi)含子。
④真核生物能夠有序地根據(jù)生長(zhǎng)發(fā)育階段的需要進(jìn)行DNA片段重排,還能在需要時(shí)增加細(xì)胞內(nèi)某些基因的拷貝數(shù)。
⑤在真核生物中,基因轉(zhuǎn)錄的調(diào)節(jié)區(qū)相對(duì)較大,它們可能遠(yuǎn)離啟動(dòng)子達(dá)幾百個(gè)甚至上千個(gè)堿基對(duì),這些調(diào)節(jié)區(qū)一般通過(guò)改變整個(gè)所控制基因5'上游區(qū)DNA構(gòu)型來(lái)影響它與RNA聚合酶的結(jié)合能力。在原核生物中,轉(zhuǎn)錄的調(diào)節(jié)區(qū)都很小,大都位于啟動(dòng)子上游不遠(yuǎn)處,調(diào)控蛋白結(jié)合到調(diào)節(jié)位點(diǎn)上可直接促進(jìn)或抑RNA聚合酶與它的結(jié)合。
⑥真核生物的RNA在細(xì)胞核中合成,只有經(jīng)轉(zhuǎn)運(yùn)穿過(guò)核膜,到達(dá)細(xì)胞質(zhì)后,才能被翻譯成蛋白質(zhì),原核生物中不存在這樣嚴(yán)格的空間間隔。
⑦許多真核生物的基因只有經(jīng)過(guò)復(fù)雜的成熟和剪接過(guò)程(maturation and splicing),才能順利地翻譯成蛋白質(zhì)。
二、真核基因的啟動(dòng)子
啟動(dòng)子是一段特定的直接與RNA聚合酶及其轉(zhuǎn)錄因子相結(jié)合、決定基因轉(zhuǎn)錄起始與否的DNA序列。不同的啟動(dòng)子對(duì)RNA聚合酶的親和力不同,所結(jié)合的反式作用因子(trans-acting factors)也不同,因此,基因轉(zhuǎn)錄活性也很不相同。
Ⅰ.典型的原核啟動(dòng)子有四大要素
轉(zhuǎn)錄起始位點(diǎn),-10區(qū),-35區(qū)和-10區(qū)與-35區(qū)之間的間隔。
原核基因轉(zhuǎn)錄起始位點(diǎn)通常是嘌呤,其序列為CAT(A為起始位點(diǎn))。
-10區(qū)是一個(gè)6堿基保守序列(常以-10為中心)。T80A95t45A60A50T96,有助于DNA的解鏈。
-35區(qū)是另一個(gè)6堿基保守序列(常以-35為中心)。T82T84G78A65C54a45
研究表明,當(dāng)-10區(qū)和-35區(qū)中心位置相距16-18bp時(shí),該啟動(dòng)子的功能較強(qiáng);相距較近或較遠(yuǎn)時(shí),起始轉(zhuǎn)錄的功能都相應(yīng)減弱。
Ⅱ.真核基因啟動(dòng)子
真核生物有3類RNA聚合酶,負(fù)責(zé)轉(zhuǎn)錄3類不同的啟動(dòng)子。
由RNA聚合酶I負(fù)責(zé)轉(zhuǎn)錄的rRNA基因,啟動(dòng)子(I類)比較單一,由轉(zhuǎn)錄起始位點(diǎn)附近的兩部分序列構(gòu)成。部分是核心啟動(dòng)子(core promoter),由-45—+20位核苷酸組成,單獨(dú)存在時(shí)就足以起始轉(zhuǎn)錄。另一部分由-170—-107位序列組成,稱為上游調(diào)控元件,能有效地增強(qiáng)轉(zhuǎn)錄效率。
由RNA聚合酶Ⅲ負(fù)責(zé)轉(zhuǎn)錄的是5SrRNA、tRNA和某些核內(nèi)小分子RNA(snRNA),其啟動(dòng)子(Ⅲ類)組成較復(fù)雜,又可被分為三個(gè)亞類。兩類5S rRNA和tRNA基因的啟動(dòng)子是內(nèi)部啟動(dòng)子(internal promoter),位于轉(zhuǎn)錄起始位點(diǎn)的下游,都由兩部分組成。第三類啟動(dòng)子由三個(gè)部分組成,位于轉(zhuǎn)錄起始位點(diǎn)上游。

由RNA聚合酶II負(fù)責(zé)轉(zhuǎn)錄的II類基因包括所有蛋白質(zhì)編碼基因和部分snRNA基因,后者的啟動(dòng)子結(jié)構(gòu)與III類基因啟動(dòng)子中的第三種類型相似,編碼蛋白質(zhì)的II類基因啟動(dòng)子在結(jié)構(gòu)上有共同的保守序列。
轉(zhuǎn)錄起始位點(diǎn)沒(méi)有廣泛的序列同源性,但個(gè)堿基為腺嘌呤,而兩側(cè)是嘧啶堿基。這個(gè)區(qū)域被稱為起始子(initiator, Inr),序列可表示為Py2CAPy5。Inr元件位于-3—+5。僅由Inr元件組成的啟動(dòng)子是具有可被RNA聚合酶II識(shí)別的最簡(jiǎn)單啟動(dòng)子形式。
多數(shù)II類啟動(dòng)子有一個(gè)被稱為TATA盒的共有序列,通常處于-30區(qū),相對(duì)于轉(zhuǎn)錄起始位點(diǎn)的位置比較固定。
三、增強(qiáng)子及其對(duì)轉(zhuǎn)錄的影響
增強(qiáng)子是指能使和它連鎖的基因轉(zhuǎn)錄頻率明顯增加的DNA序列。作為基因表達(dá)的重要調(diào)節(jié)元件,增強(qiáng)子通常具有下列性質(zhì):
1、增強(qiáng)效應(yīng)十分明顯,一般能使基因轉(zhuǎn)錄頻率增加10-200倍。經(jīng)人巨大細(xì)胞病毒增強(qiáng)子增強(qiáng)后的珠蛋白基因表達(dá)頻率比該基因正常轉(zhuǎn)錄高600-1000倍!
2、增強(qiáng)效應(yīng)與其位置和取向無(wú)關(guān),不論增強(qiáng)子以什么方向排列(5'→3'或3'→5'),甚至和基因相距3 kp,或在基因下游,均表現(xiàn)出增強(qiáng)效應(yīng);
3、大多為重復(fù)序列,一般長(zhǎng)約50bp,適合與某些蛋白因子結(jié)合。其內(nèi)部常含有一個(gè)核心序列:(G)TGGA/TA/TA/T(G),是產(chǎn)生增強(qiáng)效應(yīng)時(shí)所必需的;
4、增強(qiáng)效應(yīng)有嚴(yán)密的組織和細(xì)胞特異性,說(shuō)明只有特定的蛋白質(zhì)(轉(zhuǎn)錄因子)參與才能發(fā)揮其功能;
5、沒(méi)有基因?qū)R恍裕梢栽诓煌幕蚪M合上表現(xiàn)增強(qiáng)效應(yīng);
6、許多增強(qiáng)子還受外部信號(hào)的調(diào)控,如金屬硫蛋白的基因啟動(dòng)區(qū)上游所帶的增強(qiáng)子,就可以對(duì)環(huán)境中的鋅、鎘濃度做出反應(yīng)。
增強(qiáng)子的作用原理是什么呢?一種觀點(diǎn)認(rèn)為,增強(qiáng)子為轉(zhuǎn)錄因子提供進(jìn)入啟動(dòng)子區(qū)的位點(diǎn)。第二種認(rèn)為,增強(qiáng)子能改變?nèi)旧|(zhì)的構(gòu)象。因?yàn)樵鰪?qiáng)子區(qū)域容易發(fā)生從B-DNA到Z-DNA的構(gòu)象變化。增強(qiáng)子的功能是可以累加的。SV40增強(qiáng)子序列可以被分為兩半,每一半序列本身作為增強(qiáng)子功能很弱,但合在一起,即使其中間插入一些別的序列,仍然是一個(gè)有效的增強(qiáng)子。因此,要使一個(gè)增強(qiáng)子失活必須在多個(gè)位點(diǎn)上造成突變。對(duì)SV40增強(qiáng)子而言,沒(méi)有任何單個(gè)的突變可以使其活力降低10倍。
四、反式作用因子對(duì)轉(zhuǎn)錄的影響
真核生物啟動(dòng)子和增強(qiáng)子是由若干可以區(qū)分的DNA序列組成的,由于它們和特定的功能基因連鎖在一起,因此稱為順式作用元件。真核生物轉(zhuǎn)錄調(diào)控大多是通過(guò)順式作用元件和反式作用因子復(fù)雜的相互作用而實(shí)現(xiàn)的,下面介紹的是與順式作用成分專一性結(jié)合的一些轉(zhuǎn)錄因子。一般認(rèn)為,如果某個(gè)蛋白是體外轉(zhuǎn)錄系統(tǒng)中起始RNA合成所必需的,它就是轉(zhuǎn)錄復(fù)合體的一部分。根據(jù)各個(gè)蛋白質(zhì)成分在轉(zhuǎn)錄中的作用,能將整個(gè)復(fù)合體分為3部分:
①參與所有或某些轉(zhuǎn)錄階段的RNA聚合酶亞基,不具有基因特異性。
②與轉(zhuǎn)錄的起始或終止有關(guān)的輔助因子,也不具有基因特異性。
③ 與特異調(diào)控序列結(jié)合的轉(zhuǎn)錄因子。它們中有些被認(rèn)為是轉(zhuǎn)錄復(fù)合體的一部分,因?yàn)樗谢虼蟛糠只虻膯?dòng)子區(qū)含有這一特異序列,如TATA區(qū)和TFIID,更多的則是基因或啟動(dòng)子特異性結(jié)合調(diào)控蛋白,它們是起始某個(gè)(類)基因轉(zhuǎn)錄所必需的。
1.DNA結(jié)合結(jié)構(gòu)域基序
a. Helix-turn-helix(螺旋-轉(zhuǎn)角-螺旋)。是最早發(fā)現(xiàn)于原核生物中的一個(gè)關(guān)鍵因子,該結(jié)構(gòu)域長(zhǎng)約20個(gè)aa,主要是兩個(gè)α-螺旋區(qū)和將其隔開(kāi)的β轉(zhuǎn)角。其中的一個(gè)被稱為識(shí)別螺旋區(qū),因?yàn)樗3в袛?shù)個(gè)直接與DNA序列相識(shí)別的氨基酸。
b. Zinc finger(鋅指)。長(zhǎng)約30個(gè)aa,其中4個(gè)氨基酸(Cys或2個(gè)Cys,兩個(gè)His)與一個(gè)Zinc原子相結(jié)合。與Zinc結(jié)合后鋅指結(jié)構(gòu)較穩(wěn)定。
c. Homeodomain(同源域),最早來(lái)自控制軀體發(fā)育的基因,長(zhǎng)約60個(gè)氨基酸,其中的DNA結(jié)合區(qū)與 helix-turn-helix motif相似,人們把該DNA序列稱為homeobox。主要與DNA大溝相結(jié)合。
d. Leucine zippers(亮氨酸拉鏈)
是親脂性(amphipathic)的α螺旋,包含有許多集中在螺旋一邊的疏水氨基酸,兩條多肽鏈以此形成二聚體。每隔6個(gè)殘基出現(xiàn)一個(gè)亮氨酸。由賴氨酸(Lys)和精氨酸(Arg)組成DNA結(jié)合區(qū)。
e.堿性螺旋--環(huán)--螺旋(basic helix-loop-helix)
該調(diào)控區(qū)長(zhǎng)約50個(gè)aa殘基,同時(shí)具有DNA結(jié)合和形成蛋白質(zhì)二聚體的功能,其主要特點(diǎn)是可形成兩個(gè)親脂性α-螺旋,兩個(gè)螺旋之間由環(huán)狀結(jié)構(gòu)相連,其DNA結(jié)合功能是由一個(gè)較短的富堿性氨基酸區(qū)所決定的。
 關(guān)注我們
關(guān)注我們