LUYOR-3109高強度紫外催化光源促銷
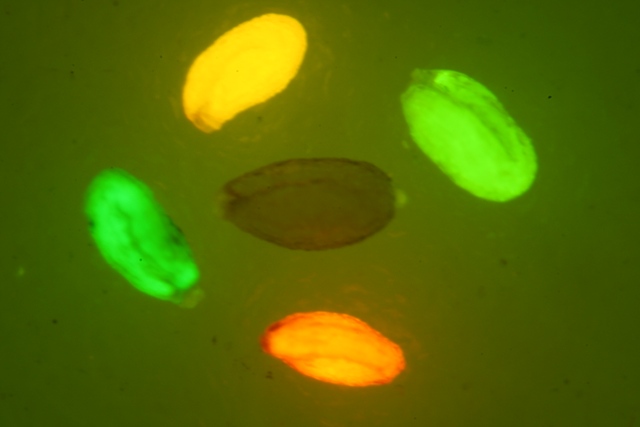
LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學(xué)透鏡,輸出紫外線強度高,...
2024-08-08


LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學(xué)透鏡,輸出紫外線強度高,...
2024-08-08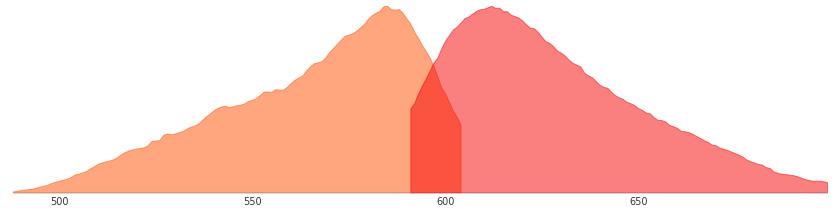
KillerRed是個完全由基因編碼的光毒性紅色熒光蛋白,可接受綠色光照(540~580nm)生成活...
2024-04-29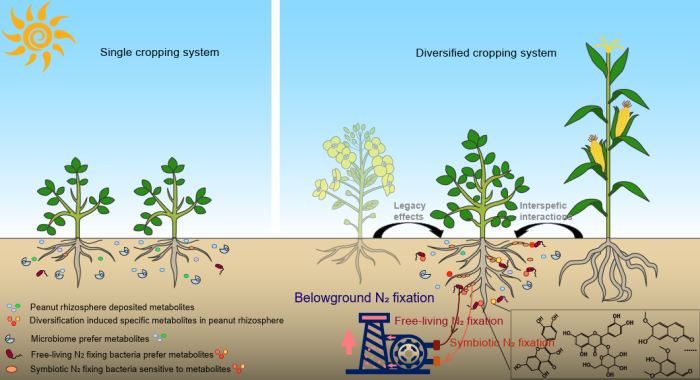
中國科學(xué)院南京土壤研究所彭新華研究員團隊陳晏副研究員在農(nóng)田長期多樣化種植下,種間植物根際對話調(diào)控土壤...
2024-04-07咨詢熱線
021-64195798作者:生命科學(xué)事業(yè)部時間:2019-11-19 08:32:30瀏覽3791 次
從分子水平研究生物大分子的結(jié)構(gòu)與功能從而闡明生命現(xiàn)象本質(zhì)的科學(xué)。自20世紀50年代以來,分子生物學(xué)是生物學(xué)的前沿與生長點,其主要研究領(lǐng)域包括蛋白質(zhì)體系、蛋白質(zhì)-核酸體系 (中心是分子遺傳學(xué))和蛋白質(zhì)-脂質(zhì)體系(即生物膜)。1953年沃森、克里克提出DNA分子的雙螺旋結(jié)構(gòu)模型是分子生物學(xué)誕生的標志。
遺傳學(xué)、基因組學(xué)和植物分子生物學(xué),是如何發(fā)展至今的?
一、列文虎克、施萊登與孟德爾
遺傳的故事都要從孟德爾開始講起!
四五百年來,植物的整理和探索。明清時代,李時珍撰寫《本草綱目》收錄藥物1892種、瑞典林奈撰寫《自然系統(tǒng)》,創(chuàng)立綱目科屬種的分析系統(tǒng)。技術(shù)方面,英國羅伯特胡克制成顯微鏡,撰寫《顯微術(shù)》,并命名了次觀察到木塞細胞(死細胞)。胡克的研究主要集中在力學(xué)和光學(xué)方面,與牛頓是同時代的人。荷蘭的眼鏡工匠師列文虎克磨制出凸透鏡適當?shù)呐浜掀饋恚箍梢詫⑽矬w放大200~300倍。1674年,列文虎克發(fā)明了世界上臺光學(xué)顯微鏡,很快利用這臺顯微鏡觀察到了血紅細胞(活細胞),證明毛細血管層的存在,描述了昆蟲、狗和人的精細胞。他也是個用顯微鏡看到細菌和原生動物的人。除了上述,描述生物學(xué)理論和技術(shù)的發(fā)展;與此同時,比利時醫(yī)學(xué)與化學(xué)家海爾蒙特,與伽利略同時代,繼承伽利略等人的思想,做了柳樹實驗,開啟植物光合作用的實驗生物學(xué)研究。
系統(tǒng)分類、光學(xué)顯微鏡、細胞、微生物、植物光合作用等詞由此而生。
時間跨過1800年,德國植物學(xué)家施萊登,與達爾文同時代,受到做植物生理學(xué)研究叔父的影響,投入并繼續(xù)使用顯微鏡和各種實驗技術(shù),開始植物胚胎學(xué)方面的深入研究。這對施萊登一生的科學(xué)活動起了決定性影響。施萊登根據(jù)他多年在顯微鏡下觀察植物組織結(jié)構(gòu)的結(jié)果,認為在任何植物體中,細胞是結(jié)構(gòu)的基本成分;低等植物由單個細胞構(gòu)成,高等植物則由許多細胞組成。發(fā)表論文《論開花植物胚株的發(fā)育史》首篇論文,《植物發(fā)生論》研究了植物細胞的形成,在這個基礎(chǔ)上,施萊登提出了植物細胞學(xué)說。又撰寫《植物學(xué)概論》、《植物及其生活》等書籍。與此同時,英國達爾文完成環(huán)球航行,撰寫了《物種起源》一書,提出生物進化學(xué)說,書中達爾文提到了遺傳原材料是變異。并記錄了在航行船上觀察到的植物向光生長現(xiàn)象,撰寫《植物的運動力》。
細胞、植物學(xué)、植物發(fā)生、細胞發(fā)生、遺傳、變異、植物向光性等詞由此而生。
時間進入1865年孟德爾的時代,奧地利遺傳學(xué)家孟德爾,與達爾文同時代,受到園藝家庭背景的影響,進入維也納大學(xué)深造,受到相當系統(tǒng)和嚴格的科學(xué)教育和訓(xùn)練。孟德爾受到德國統(tǒng)計學(xué)家恩格爾(同時是細胞理論的推動者)的影響,與自己長期思索,認識到理解那些使遺傳性狀代代恒定的機制更為重要。1856年,從維也納大學(xué)回到修道院不久,孟德爾就開始了長達8年的豌豆實驗。孟德爾使用統(tǒng)計學(xué)理論、豌豆人工培植技術(shù)和大量的實驗,發(fā)現(xiàn)了生物遺傳的基本規(guī)律,并得到了相應(yīng)的數(shù)學(xué)關(guān)系式。人們分別稱他的發(fā)現(xiàn)為“孟德爾定律”(即孟德爾遺傳分離規(guī)律)和“孟德爾第二定律”(即基因自由組合規(guī)律),它們揭示了生物遺傳奧秘的基本規(guī)律。
遺傳、遺傳學(xué)、性狀、表型、遺傳因子等詞由此而生。

二、薩頓與摩爾根
直到1900年孟德爾逝世16年后,他的遺傳學(xué)說才又被人們重新發(fā)現(xiàn)。3:1的性狀分離比的遺傳現(xiàn)象引起了的注意,但是更好的解釋還是孟德爾遺傳定律。美國遺傳學(xué)家沃爾特薩頓,在導(dǎo)師威爾遜已經(jīng)提出“每一個生命科學(xué)問題的答案都必須在細胞中尋找”的論斷后,率先在1902年觀察發(fā)現(xiàn)細胞中存在成對的染色體成,指出染色體就是孟德爾在其工作中假設(shè)的遺傳因子(孟德爾的工作被重新發(fā)現(xiàn)才兩年)。1903年,他在另一篇論文中堅持認為:染色體帶有基因;每個性細胞只具有一對染色體,具有哪是一對是隨機決定的。并將遺傳因子改名為基因。
接著是時間進入摩爾根的時代,美國遺傳學(xué)家摩爾根,由于長期研究果蠅對孟德爾遺傳定律存有質(zhì)疑。他認為這些定律可能只適合于豌豆而不適用于其他生物。摩爾根著手建立了果蠅實驗系統(tǒng),研究果蠅的性狀到底是怎樣遺傳的。當子二代紅、白果蠅的比例正好是3:1時,摩爾根對孟德爾更加佩服了。摩爾根確信,染色體就是基因的載體!摩爾根還推算出了各種基因的染色體上的位置,繪制果蠅的4對染色體上的基因所排列的位置圖,基因?qū)W說從此誕生。1926年撰寫了《基因論》,對基因這一遺傳學(xué)基本概念進行了具體而明確的描述。
摩爾根進一步提出,染色體上基因存在連鎖群現(xiàn)象,但并不像鐵鏈一樣牢靠,有時染色體也會發(fā)生斷裂,與另一條染色體互換部分基因。基因在染色體上的位置距離越遠,它們之間出現(xiàn)染色體交換基因的頻率就越大。摩爾根在《科學(xué)》雜志上發(fā)表一篇闡述連鎖與互換定律的理論文章,“連鎖與互換定律”是摩爾根在遺傳學(xué)領(lǐng)域的一大貢獻,它和孟德爾的分離定律、自由組合定律一道,被稱為遺傳學(xué)三大定律。
染色體、遺傳學(xué)、連鎖互換、基因突變重組、遺傳連鎖圖、物理圖譜等詞由此出現(xiàn)。

三、德爾布呂克、鮑林、沃森與克里克
1928年,英國醫(yī)生格里菲斯發(fā)現(xiàn)并記錄了肺炎雙球菌遺傳轉(zhuǎn)化實驗,即可是了遺傳轉(zhuǎn)化的研究,也開始了科學(xué)界的次大探討,遺傳物質(zhì)是蛋白質(zhì)還是DNA?加拿大化學(xué)家艾弗里率先純化核酸,并驗證DNA是遺傳物質(zhì),而不是蛋白質(zhì)。德國物理學(xué)家德爾布呂克(與李比希是親戚)采用最簡單的生物來探討“基因的化學(xué)本質(zhì)是什么”的問題。德爾布呂克與摩爾根關(guān)系甚密,但是研究的果蠅使他感到一籌莫展,因為他認為果蠅過于復(fù)雜而不適應(yīng)于物理學(xué)家慣有的簡單性思維。德爾布呂克選擇噬菌體作為自己做研究的材料,很快和赫爾希、盧利亞、蔡斯等發(fā)現(xiàn)噬菌體以DNA核酸的方式,遺傳轉(zhuǎn)化侵染了大腸桿菌,證明DNA是遺傳物質(zhì)。
噬菌體、大腸桿菌、模式生物等詞由此出現(xiàn)。
美國化學(xué)家鮑林,鮑林在學(xué)習(xí)了量子力學(xué)后,率先將量子力學(xué)應(yīng)用于化學(xué)鍵的研究中,得出關(guān)于有機化合物分子結(jié)構(gòu)的“共振論”,并且成功解析蛋白質(zhì)的a螺旋結(jié)構(gòu)模型。1951年鮑林結(jié)合他在血紅蛋白進行的實驗研究,以及對肽鏈和肽平面化學(xué)結(jié)構(gòu)的理論研究,提出了α螺旋和β折疊是蛋白質(zhì)二級結(jié)構(gòu)的基本構(gòu)建單元的理論。同時鮑林也是首先想到x-射線衍射晶體結(jié)構(gòu)測試法,引入到蛋白質(zhì)結(jié)構(gòu)測定的人,并且推導(dǎo)了衍射圖譜計算蛋白質(zhì)中原子坐標的公式。鮑林進一步有投入到DNA分子結(jié)構(gòu)解析的工作上來。
英國卡文迪許(研究空氣和引力常數(shù),與牛頓同時代)實驗室的結(jié)構(gòu)生物學(xué)派的威爾金斯和富蘭克林,在布拉格父子使用X射線研究晶體結(jié)構(gòu)工作的基礎(chǔ)上,研究了手關(guān)于DNA分子結(jié)構(gòu)的圖樣。英國物理家克里克和年輕的美國生物學(xué)者沃森,基于合理的猜想和生物學(xué)知識,于1953年率先解析DNA分子的雙螺旋結(jié)構(gòu)。終于將基因研究推向分子水平,推動了分子生物學(xué)的誕生。DNA雙螺旋結(jié)構(gòu)的發(fā)現(xiàn),進一步促進了DNA半保留復(fù)制方式的發(fā)現(xiàn)和遺傳密碼的研究工作。1961年,遺傳密碼的破譯使得中心法則趨于完善。留下了更多的問題是,基因組序列的破譯工作。
結(jié)構(gòu)生物學(xué)、分子生物學(xué)等詞由此出現(xiàn)。
四、雅各布、莫諾德、萊夫科維茨與克比爾卡
1961年,雅各布和莫諾德提出的乳糖操縱子學(xué)說,詳細研究了大腸桿菌分解乳糖的操縱子模型。發(fā)現(xiàn)基因表達調(diào)控的阻遏機制和模型。提示,基因通常是在阻遏條件下表達的,基因的表達被阻遏蛋白阻遏。在外界信號改變時,信號刺激蛋白質(zhì)活性的改變進而調(diào)劑基因的表達。
萊夫科維次與科比爾卡,1968年,利用放射學(xué)方法追蹤細胞受體變化。與科比可卡揭示了G蛋白偶聯(lián)受體,這一大類受體的工作模式。蛋白質(zhì)相互作用的和受體的尋找等分子水平的工作也慢慢開始。1978年,下村修和約翰森在深海水母中發(fā)下GFP熒光蛋白,有力的推進了生命科學(xué)行業(yè)的發(fā)展。
受體、信號通路、蛋白質(zhì)相互作用、激酶、轉(zhuǎn)錄因子、阻遏蛋白、正負調(diào)控等詞由此出現(xiàn)。
五、史密斯、伯格、菲爾與海洛
1972年,史密斯和伯格等率先使用限制性核酸內(nèi)切酶和DNA連接酶,實現(xiàn)了對DNA的人工操作和轉(zhuǎn)基因操作。他們利用轉(zhuǎn)基因技術(shù)所注冊的生物技術(shù)公司,生產(chǎn)了市場上各種蛋白類藥物,如胰島素和人類生長激素等,市值達到3000億元。1992年,菲兒和海洛在微生物中發(fā)現(xiàn)以RNA干擾的方式控制基因信息流動的機制,進而促進了RNAi基因沉默編輯技術(shù)的發(fā)展。2013年,劉大衛(wèi)和張峰等人在微生物中發(fā)現(xiàn)以CRSPR/Cas9的方式控制基因信息流動的機制,直接促進了基因編輯技術(shù)的發(fā)展。
轉(zhuǎn)基因技術(shù)、基因工程、基因編輯技術(shù)等詞由此出現(xiàn)。
")
六、Holliday、Fischer和Krebs
1987年,Holliday提出表觀遺傳概念,認為DNA甲基化修飾水平的改變,可以引起基因表達活性的改變。Fischer和Krebs對蛋白質(zhì)可逆磷酸化調(diào)節(jié)糖代謝過程的研究揭開了蛋白在表觀修飾的研究。現(xiàn)代表觀遺傳學(xué)認為,在基因的DNA序列不發(fā)生改變的情況下,基因的表達水平與功能發(fā)生改變,可以產(chǎn)生可以遺傳的表型。主要的表觀遺傳標記存在于染色體的不同水平,包括DNA和組蛋白修飾、組蛋白多樣性、直接結(jié)合與DNA或組蛋白上的染色體上的非組蛋白修飾等。
表觀遺傳學(xué)、甲基化、磷酸化、乙酰化等詞由此出現(xiàn)。
七、Sanger、穆利斯和田中耕一
桑格在完成蛋白質(zhì)測序任務(wù)后,1975年又開發(fā)了雙脫氧核苷酸末端終止測序法漂亮的完成了DNA測序任務(wù)。并測完了噬菌體的基因組。隨后各種基因組計劃開始了,直到2003年人類基因組也順利測完,這意味著,測序工作進入高速發(fā)展階段,更加趨向于變成一種試用的工具。1983年,穆利斯發(fā)現(xiàn)了DNA的體外擴增技術(shù)PCR,使得基因操作變得更加便捷,分子生物的工具更加豐富和了,極大地推動全基因組測序工作的進展。與此同時,日本島津儀器公司研發(fā)工程師田中耕一,在使用質(zhì)譜儀解析生物大分子結(jié)構(gòu)的技術(shù)改進工作中發(fā)揮了突出的貢獻。終于使得蛋白質(zhì)組學(xué)研究,成為可能。
基因組、轉(zhuǎn)錄組、測序、蛋白質(zhì)組、代謝組、單細胞測序等詞由此出現(xiàn)。
 關(guān)注我們
關(guān)注我們