LUYOR-3109高強(qiáng)度紫外催化光源促銷
LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學(xué)透鏡,輸出紫外線強(qiáng)度高,...
2024-08-08


LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學(xué)透鏡,輸出紫外線強(qiáng)度高,...
2024-08-08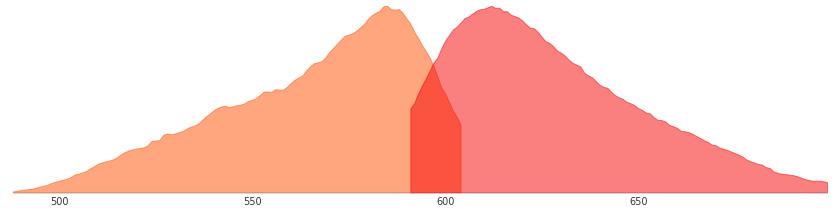
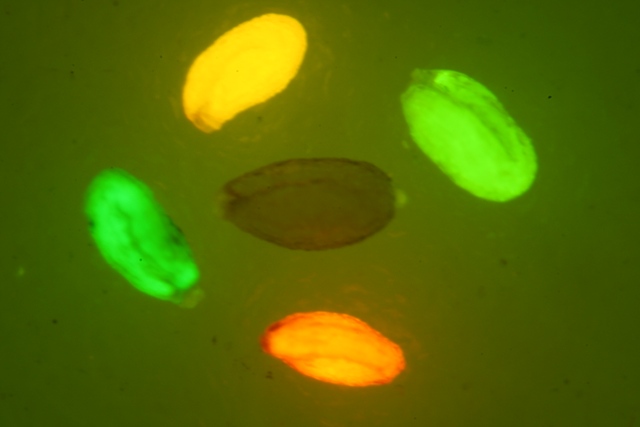
KillerRed是個(gè)完全由基因編碼的光毒性紅色熒光蛋白,可接受綠色光照(540~580nm)生成活...
2024-04-29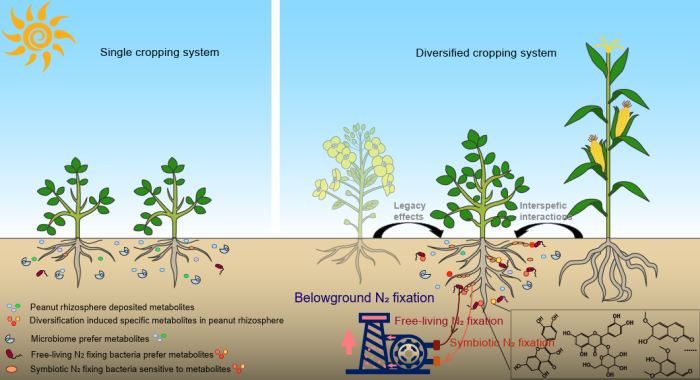
中國(guó)科學(xué)院南京土壤研究所彭新華研究員團(tuán)隊(duì)陳晏副研究員在農(nóng)田長(zhǎng)期多樣化種植下,種間植物根際對(duì)話調(diào)控土壤...
2024-04-07咨詢熱線
021-64195798作者:生命科學(xué)事業(yè)部時(shí)間:2019-12-14 20:27:46瀏覽17252 次
DNA甲基化是最早發(fā)現(xiàn)的修飾途徑之一,可能存在于所有高等生物中。DNA甲基化能關(guān)閉某些基因的活性,去甲基化則誘導(dǎo)了基因的重新活化和表達(dá)。
DNA甲基化與基因表達(dá)
DNA甲基化是最早發(fā)現(xiàn)的修飾途徑之一,可能存在于所有高等生物中。DNA甲基化能關(guān)閉某些基因的活性,去甲基化則誘導(dǎo)了基因的重新活化和表達(dá)。
1.DNA甲基化的主要形式
5-甲基胞嘧啶,N6-甲基腺嘌呤和7-甲基鳥(niǎo)嘌呤。在真核生物中,5-甲基胞嘧啶主要出現(xiàn)在CpG和CpXpG中,原核生物中CCA/TGG和GATC也常被甲基化。
真核生物細(xì)胞內(nèi)存在兩種甲基化酶活性:一種被稱為日常型(mainte-nance)甲基轉(zhuǎn)移酶,另一種是兩種甲基化酶從頭合成(denovo synthesis)甲基轉(zhuǎn)移酶。前者主要在甲基化母鏈(模板鏈)指導(dǎo)下使處于半甲基化的DNA雙鏈分子上與甲基胞嘧啶相對(duì)應(yīng)的胞嘧啶甲基化。日常型甲基轉(zhuǎn)移酶常常與DNA內(nèi)切酶活性相耦聯(lián),有3種類型。II類酶活性包括內(nèi)切酶和甲基化酶兩種成分,而I類和III類都是雙功能酶,既能將半甲基化DNA甲基化,又能降解外源無(wú)甲基化DNA。
由于甲基化胞嘧啶極易在進(jìn)化中丟失,所以,高等真核生物中CG序列遠(yuǎn)遠(yuǎn)低于其理論值。哺乳類基因組中約存在4萬(wàn)個(gè)CG islands,大多位于轉(zhuǎn)錄單元的5'區(qū)。

沒(méi)有甲基化的胞嘧啶發(fā)生脫氨基作用,就可能被氧化成為U,被DNA修復(fù)系統(tǒng)所識(shí)別和切除,恢復(fù)成C。已經(jīng)甲基化的胞嘧啶發(fā)生脫氨基作用, 它就變?yōu)門(mén), 無(wú)法被區(qū)分。因此, CpG序列極易丟失。
二、 蛋白質(zhì)磷酸化與基因表達(dá)
蛋白質(zhì)的磷酸化反應(yīng)是指通過(guò)酶促反應(yīng)把磷酸基團(tuán)從一個(gè)化合物轉(zhuǎn)移到另一個(gè)化合物上的過(guò)程,是生物體內(nèi)存在的一種普遍的調(diào)節(jié)方式,在細(xì)胞信號(hào)的傳遞過(guò)程中占有極其重要的地位。

已經(jīng)發(fā)現(xiàn)在人體內(nèi)有多達(dá)2000個(gè)左右的蛋白質(zhì)激酶和1000個(gè)左右的蛋白質(zhì)磷酸酶基因。蛋白質(zhì)的磷酸化是指由蛋白質(zhì)激酶催化的把ATP或GTP上γ位的磷酸基轉(zhuǎn)移到底物蛋白質(zhì)氨基酸殘基上的過(guò)程,其逆轉(zhuǎn)過(guò)程是由蛋白質(zhì)磷酸酶催化的,稱為蛋白質(zhì)脫磷酸化。
1. 蛋白質(zhì)磷酸化在細(xì)胞信號(hào)轉(zhuǎn)導(dǎo)中的作用
(1). 在胞內(nèi)介導(dǎo)胞外信號(hào)時(shí)具有專一應(yīng)答特點(diǎn)。與信號(hào)傳遞有關(guān)的蛋白激酶類主要受控于胞內(nèi)信使,如cAMP,Ca2+,DG(二酰甘油,diacyl glycerol)等,這種共價(jià)修飾調(diào)節(jié)方式顯然比變構(gòu)調(diào)節(jié)較少受胞內(nèi)代謝產(chǎn)物的影響。
(2).蛋白質(zhì)的磷酸化與脫磷酸化控制了細(xì)胞內(nèi)已有的酶"活性"。與酶的重新合成及分解相比,這種方式能對(duì)外界刺激做出更迅速的反應(yīng)。
(3).對(duì)外界信號(hào)具有級(jí)聯(lián)放大作用;
(4).蛋白質(zhì)的磷酸化與脫磷酸化保證了細(xì)胞對(duì)外界信號(hào)的持續(xù)反應(yīng)。
被磷酸化的主要氨基酸殘基:絲氨酸、蘇氨酸和酪氨酸。組氨酸和賴氨酸殘基也可能被磷酸化。
三、基因重排的分子機(jī)制
早在50年代,人們就認(rèn)識(shí)到抗體分子的每一條鏈都是由高度多變的V區(qū)和相對(duì)不變的C區(qū)組成的,V區(qū)賦予抗體分子對(duì)抗原的特異性。抗體分子V區(qū)的多樣性和C區(qū)的穩(wěn)定性顯然是矛盾的。Dreyer和 Bennett 于1965年提出假設(shè),認(rèn)為每條抗體鏈實(shí)際上至少由兩個(gè)基因所編碼,其中一個(gè)是恒定的,一個(gè)是可變的。
1983年,Tonegawa (Nature, 302:575-581)在對(duì)產(chǎn)生抗體的骨髓瘤及漿細(xì)胞瘤進(jìn)行研究時(shí)發(fā)現(xiàn),產(chǎn)生抗體的細(xì)胞中Ig基因結(jié)構(gòu)與其它不合成抗體分子的細(xì)胞中的結(jié)構(gòu)不一樣。在所有物種中,胚系Ig基因的構(gòu)成基本上相同。Ig重鏈和輕鏈(λ和κ鏈)基因座都由多個(gè)編碼V區(qū)和C區(qū)蛋白質(zhì)的基因組成,并被非編碼的DNA所分隔。抗體分子由4條(兩對(duì))多肽鏈組成,包括兩條相同的輕鏈(L-chain)和兩條相同的重鏈(H-chain)。輕鏈和重鏈在相對(duì)分子質(zhì)量上有較大差別,前者約2.3x104,后者則介于 5.3x104-7.0x104之間。 所有Ig分子都含有兩類輕鏈中的一類,即κ型或λ型。Κ型和λ型輕鏈的恒定區(qū)和可變區(qū)的氨基酸序列是不同的。在小鼠中,95%的抗體輕鏈?zhǔn)铅市停祟惪贵w輕鏈中,κ型和λ型各占50%左右。免疫球蛋白重鏈基因DNA重排以后,大量間隔序列被切除,使位于J-Cμ之間的增強(qiáng)子序列得以發(fā)揮作用,增強(qiáng)基因轉(zhuǎn)錄。
 關(guān)注我們
關(guān)注我們