LUYOR-3109高強度紫外催化光源促銷
LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學透鏡,輸出紫外線強度高,...
2024-08-08

作者:生命科學事業部時間:2019-11-05 09:30:48瀏覽16843 次
自1999年紅色熒光被發現后,引起了科學家廣泛的興趣,人們對其理化性質和發光機制進行了深入的研究;通過對其修飾,人們得到了聚集程度低、成熟速率更快、發光強度更強和發射波長更長的突變株,這些都極大地方便了DsRed在科學研究中的應用。目前,DsRed的使用還遠未有G FP廣泛,相信隨著對DsRed研究的深入,DsRed將成為人們進行科學研究的重要手段。
1999年,Matz 等從印度洋2太平洋地區的珊瑚蟲中分離出6種與綠色熒光蛋白(green fluorescent proteins ,GFP )同源的熒光蛋白,并鑒定了它們的光譜性質。其中來源于Discosoma sp.紅色熒光蛋白(drFP583)在紫外線的照射下可發射紅色熒光,其更大吸收波長為558nm ,更大發射波長為583nm 。其發射波長較長,靈敏度與信噪比均比GFP 高,為基于GFP 的體內研究提供了一個很好的互補工具。但是,drFP583具有自身的缺點,如寡聚化、成熟過程緩慢和對細胞有毒性等限制了它的應用。因此,人們對其進行各種修飾和改進,以得到不同發光特性、不同聚集狀態及適于不同細胞的突變體。更先進行突變實驗的是drFP583,而Clontech 公司已將它的一個低毒、低寡聚化及成熟快的突變體E572NA 商業化,商品名為DsRed [2]。隨著DsRed 的修飾和改進,其應用領域越來越廣,而且它與GFP 相結合,為分子生物學和細胞生物學等相關領域的研究提供了一個新的、快捷的檢測手段。
1.1 蛋白質的結構
DsRed 基因編碼的蛋白質由225個氨基酸組成,相對分子質量(M r )為25.9ku 。它的一級結構與GFP的同源性很低,僅為23%;但它們的二級結構很相似。DsRed 的單體由11條β桶狀結構(β2bar 2rel )繞成的一個圓柱體,一條α螺旋纏繞在圓柱體的軸位置,生色團附著在α螺旋上,包埋于圓柱體中心,這種方式被稱為β罐(β2can )。
1.2 生色團
DsRed 是Mr為26ku的多肽,其內部的G ln 2T yr 2G ly (66~68位殘基)三肽自催化形成與avG FP 相似的生色團。在生色團附近的幾個氨基酸殘基相當保守,這可能是生色團形成所必需的(如G lu225,Arg95,對應于綠色熒光的G lu222和Arg95)。它的更大吸收和發射波長分別為558和583nm (在494和280nm 處分別有一個小峰)。
Baird 等研究了DsRed 的成熟動力學,發現室溫下該蛋白更先形成綠色熒光生色團(更大吸收和發射波長分別為475和499nm ),其熒光強度在更高峰可維持7h ,然后逐漸降為零,歷時2d 。與此同時,紅色熒光在27h 達到更大強度的50%,48h 后,達到更大強度的90%。對野生型的DsRed 進行隨機突變,得到了發射綠光的突變株。基于這個發現,人們推測DsRed 發色團形成的機制主要為:先形成GFP 樣的綠色陰離子發色團中間體(吸收和發射波長分別為480和500nm ),然后經過氧化反應,轉變為成熟的紅色熒光發色團(吸收和發射波長分別為558和583nm )。總之,DsRed 的發色團是GFP 發色團的延伸。實驗結果表明,DsRed 的發色團只有在寡聚化的情況下才能形成及發揮作用。對DsRed 堿變性、酸變性及胍變性實驗表明,DsRed 的發色團要經過幾步折疊才可形成,而寡聚化在它的成熟過程中起重要的作用。
1.3 DsRed的優缺點
與GFP相比,DsRed的激發和發射波長較長,其發射峰位于培養基、組織培養器材及細胞成分等產生的熒光背景范圍之外,具有較高的信噪比;而且在細胞內熒光轉換效率高,更易檢測。DsRed具有很高的消光系數和熒光量子產量,這些特性表明,DsRed發射熒光的強度要比羅丹明B等染料和更好的GFP突變體的強度高得多;其抗漂白能力強于GFP及其黃色熒光突變株,以及增強型綠色熒光蛋白(EG FP)和增強型黃色熒光蛋白(ECFP)。另外,GFP的諸多突變體在酸性環境下很容易發生熒光淬滅,且在很多條件下極易生成有害物質;而DsRed對pH值不敏感,pH值為4.5~12時仍保持穩定,這使其使用范圍更加廣泛。目前,人們正在研究DsRed對pH不敏感的原因,并將其應用在GFP的優化方面。DsRed更大的缺點是緩慢和不完全的成熟過程及寡聚化。緩慢的成熟過程阻止了它作為報告基因的一些應用,如檢測短期的基因表達及生物體快速發育時的一些情況。對GFP進行突變可獲得快速成熟的突變株,人們也對DsRed進行類似突變以期得到快速成熟的突變株。

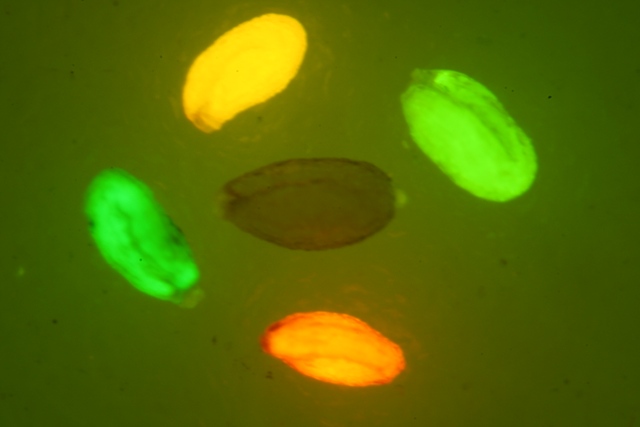
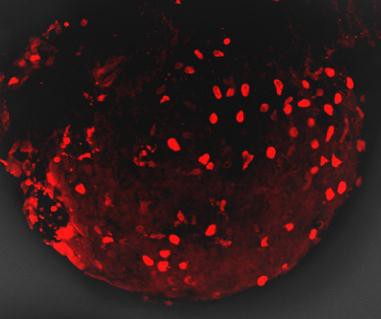
DsRed紅色熒光蛋白的激發波長為554nm,發射波長為591nm
基于對DsRed結構的理解和生色機制的闡明,人們采用隨機突變或定點突變對DsRed進行優化。與DsRed相比,其突變株的特點主要是減少寡聚化,加快成熟速率和改變波長等。
2.1 減少寡聚化
2.1.1 寡聚化的原因 利用超速離心分析法、能量共振轉移法、酵母雙雜交法分析DsRed在體內外的表達情況,結果表明,無論在體內還是在體外,DsRed 都傾向形成寡聚體,而且它需要幾天時間才能完全成熟。通過時間依賴的熒光各向異性和單分子光譜實驗,證明了DsRed即使在納摩爾級(nm ol?L-1)以下濃度時也存在寡聚體,濃度高時甚至可形成八聚體[4,6]。Yarbrough等[3]和Wall等[7]通過X射線分別得到了DsRed的晶體結構。分析了DsRed的立體結構,發現四聚體表面的疏水基團較少;用計算機分析DsRed四聚體的靜電位(勢),發現蛋白表面主要帶負電荷,而且每個單體的N端均含有幾個帶正電荷的氨基酸殘基,它們通過靜電相互作用導致了蛋白的聚集。人們推測每個DsRed四聚體通過4個鹽橋與鄰近的四聚體相互作用,形成網狀的多聚結構。4個配位鍵的靜電相互作用使得“天線樣”的結構非常穩定[2]。降低DsRed的寡聚化程度和加快它的成熟過程,可使其具有更好的應用前景,因此,有必要對DsRed進行修飾和改進。
2.1.2 減少寡聚化的方法 人們認為,N端帶正電的氨基酸殘基可能是引起DsRed聚集的主要原因。因此可通過去除熒光蛋白N端的幾個氨基酸殘基,或者用帶負電荷的或中性氨基酸取代N端的Lys和Arg殘基,來獲得非聚集的珊瑚蟲屬(Anthozoa)熒光蛋白的突變體。對DsRed進行定點突變(V105A, I161T,S197A),得到了一個在大腸桿菌中快速及較完全折疊的突變體,命名為E57;然后以E57作母體,進行再次突變。該實驗中,三個突變位點(R2A, K5E和K9T)分別以不同的組合進行氨基酸的替換;并用大腸桿菌表達這些突變體,純化后用聚丙烯酰胺凝膠電泳(PAGE)分析:與母體E57相比,這些突變體的聚集水平均有所下降,R2A突變似乎對聚集水平的下降影響更大。突變株E572NA(R2A,K5E和K9T)的體內外實驗表明,該蛋白聚集程度低,而且成熟速率是最快的;它的更大激發波長、熒光強度與母體E57相似。動物實驗表明,該蛋白的細胞毒性明顯低于DsRed和E57。現在E572NA已經被Clon2 tech公司商業化,名稱為DsRed2[2]。而Campbell 等[8]替換了DsRed序列中的33個氨基酸殘基,得到了單體紅色熒光蛋白mRFP1,該蛋白比DsRed更適于與G FP等進行多色熒光標記,但是,它仍具有熒光強度偏低等缺點,需進一步改進。
2.2 改變發射波長的突變
2.2.1 發射綠色熒光突變株
人們曾對DsRed進行隨機和定點突變,得到突變株P37S,K83R,N42H 和T217S,可將DsRed穩定在綠色中間體,更大吸收波長為480nm;另一個中間體的更大吸收波長為408nm,實驗證明它們是生色團的質子化和脫質子化形式,這與G FP的生色團類似[4,9]。這些綠色熒光突變株可作為紅色熒光生色團形成機制的證據。
2.2.2 發射波長紅移(red2shift,RS)突變株 據報道,熒光蛋白發射波長越長,其檢測靈敏度越高。人們對mRFP1進行定點突變(突變位點:T202I,R3K, S112T,P188Q,M182K和V96I),得到增強型的單體mRFPmars,其更大吸收和發射波長分別為585和602 nm。而Fradk ov等對兩個紅色熒光蛋白(dsFP593和drFP583)進行隨機突變,得到突變株Ds/drFP616,其發射波長明顯延長(616nm),但聚集性還很高,隨后對Ds/drFP616進一步突變(K5E和K9T),其突變株Ds/drFP6162K5E/K9T的聚集性明顯降低。
2.2.3 發射波長可變的熒光蛋白 最近,人們還發現了一個有趣的DsRed突變體,命名為E5,又稱“熒光計時器(fluorescent timer)”。它發射的熒光顏色隨蛋白濃度的變化而改變,從鮮綠色、黃色、橙色,最后變成紅色[12]。將這種蛋白的基因插入到幾種不同類型的細胞基因組中,使它處于某個啟動子控制之下,觀察基因開啟和轉錄等,從而使研究者方便、快速地了解到基因轉錄的適時信息。對DsRed2T imer 進行突變(R2A,K5E,K9T),突變體命名為T imer2 NA。它與母體DsRed2T imer的成熟過程及顏色變化相同,實驗證明它不形成聚集體[2]。
2.3 適合特定細胞的修飾
Sorensen等[13]利用隨機突變的方法,得到可以在細菌中高度表達的快速成熟的DsRed.T3突變株DsRed.T32S4T。用流式細胞儀檢測,DsRed.T32S4T 的發光強度足夠檢測到沙門菌基因的表達;然而,在利什曼原蟲和HeLa細胞中,看不到熒光。說明突變引起的mRNA折疊變化影響了原核的轉錄系統,但對真核轉錄系統不起作用。而將DsRed基因置于不同生物專用的啟動子下,則可以檢測該生物的轉化效率及基因表達的效率。如將DsRed啟動子置于絲狀真菌常用的啟動子Pgpd之下,得到了載體Pgpd2DsRed,用其轉化鐮刀菌,可觀察到鐮刀菌對西紅柿的侵染、分布及致病力,還可以用它與含GFP的載體共轉化青霉屬和木霉屬的真菌,得到雙標記的真菌。
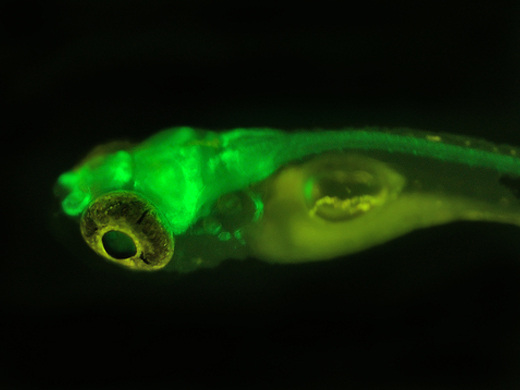
目前,DsRed已應用于啟動子分析、轉基因、蛋白質和細胞定位、藥物篩選、細胞、整個器官的雙標記和三標記培養,以及利用熒光共振能量轉移(FRET)研究蛋白與蛋白的相互作用;另外,DsRed還可作為胞內不同信使的熒光傳感器。為研究ABC轉運基因如何在線蟲中起作用,人們設計了融合有熒光蛋白的啟動子,以觀察這些基因何時在體內開啟[15];還可將DsRed與蛋白質融合,分析蛋白的表達與定位;或者構建融合有DsRed 的基因組文庫或cDNA文庫。最近,人們利用轉基因技術獲得了一個全身發射綠色熒光的小鼠,在小鼠體內移植發射紅色熒光的胰腺腫瘤,可清楚地觀察到腫瘤和基質的相互作用,這為人們研究腫瘤的發生、發展提供了很好的手段。DsRed與顯微鏡技術相結合的熒光成像技術,可應用于藥物代謝、藥劑學等學科,用來分析藥物遞送載體在組織器官、細胞中的分布和定位等。
4 結語
自1999年紅色熒光被發現后,引起了科學家廣泛的興趣,人們對其理化性質和發光機制進行了深入的研究;通過對其修飾,人們得到了聚集程度低、成熟速率更快、發光強度更強和發射波長更長的突變株,這些都極大地方便了DsRed在科學研究中的應用。目前,DsRed的使用還遠未有G FP廣泛,相信隨著對DsRed研究的深入,DsRed將成為人們進行科學研究的重要手段。
如果要觀察Dsred紅色熒光蛋白(red Fluorescent Protein)的表達,美國路陽生產的便攜式熒光蛋白激發光源可以選擇LUYOR-3260GR和LUYOR-3415(X)G系列雙波長熒光蛋白激發光源。紅色熒光蛋白采用綠光激發,佩戴LUV-50A紅色觀察眼鏡觀察,如希望提供更多詳細信息,可直接聯系上海路陽生物技術有限公司的銷售客服。
 關注我們
關注我們